I feel bad for the gene. Why? Well first, just take a look at his face.
Why is Gene so sad? I think it's because we're always talking about him behind his back, saying things like “it's genetic” and, “so-and-so has the gene for _____.” Because we decided who he was decades before we even met him. And the more we learn about him, the more we continue to talk about him as if we've learned nothing. The sadness upon his face seems to say, “How come you never listen?”
The word “gene” was introduced in 1909, but the structure of DNA was not determined until the 1950s. The word “gene” was introduced to mean, basically, a unit of biological heredity responsible for a specific heritable trait.
The idea that each trait had a specific unit of heredity behind it was of great service to the eugenics movement, as I discussed in my
introduction to this series. Soon after, it became very useful to the neo-Darwinian Modern Synthesis that unified Mendel's laws of inheritance with Darwin's theory of natural selection as the primary creative force responsible for the diversification of all life from a common ancestor.
Nevertheless, the discovery of molecular genetics, from its inception, began to add much more to the story.
No doubt genes carry heritable information. And no doubt there are variants of each gene called alleles, which contribute to variation in the traits of an organism that are subject to natural selection.
But is that all a gene does? Quite certainly not. With the discovery of DNA, molecular biology began showing that genes are designed to bestow flexibility upon an organism, not simply the ability to inherit characteristics, but the ability to change characteristics in response to the environment and in response to the organism's own needs.
The remarkable ability of the cell to access and utilize its genome, much like we use our computer programs, will be the topic of the next post in this series.
In this post, I'd like to focus on another reason that Gene might be sad. Perhaps when we assume that a heritable disease is “genetic” Gene feels unjustly blamed, and perhaps when we assume that the marvelous and beautiful functional and aesthetic traits of an organism are “genetic” Gene feels flattered but somewhat uneasy of the high expectations laid upon him. Perhaps we are being somewhat demanding in assuming that Gene should carry the burden of biological heredity all by himself.
Please forgive my anthropomorphizing for a moment, as I promise this post actually contains some science, following shortly below.
There is a type of biological inheritance that is not encoded in the DNA within the nucleus of our cells called cytoplasmic inheritance. The cytoplasm consists of the fluid that surrounds the nucleus called the cytosol and all the organelles within it. Organelles are the “little organs” of the cell.
While we inherit half of our nuclear DNA from our mother and half from our father, we inherit mostly our mother's cytoplasm. This includes our mother's mitochondria, which contains its own genome, but it also includes all the other organelles of our mother's egg cell, and these all carry heritable information that is not encoded in any DNA at all, as will be discussed further below.
Cytoplasmic inheritance is commonly discussed in genetics and molecular biology textbooks. One modern way of demonstrating cytoplasmic inheritance is to take a nucleus from one species and transfer it to a denucleated cell of another species. This technique usually fails, demonstrating the need for compatability between the nucleus and the rest of the cytoplasm, but the few success stories are quite fascinating.
Six years ago, for example, Chinese researchers successfully transplanted nuclei from a common carp genetically engineered to express human growth hormone into the egg cells of a goldfish. Remarkably, the cloned hybrid shared certain characteristics that were intermediate between the two types of fish, rather than simply exhibiting the characteristics of the carp. Here's a picture of the hybrid and its parental species:
Sun YH, Chen SP, Wang YP, Hu W, Zu ZY. Cytoplasmic Impact on Cross-Genus Cloned Fish Derived from Transgenic Common Carp (Cyprinus carpio) Nuclei and Goldfish (Carassius auratus) Enucleated Eggs. Biology of Reproduction. 2005;72:510-515.
The cloned hybrid is on the left. The hybrid's nucleus came from the carp, shown in the middle, and its cytoplasm came from the goldfish, shown on the right.
The authors say there was little difference in the appearance of the hybrid from the carp, and indeed, the hybrid looks a lot closer to the carp than to the goldfish. However,
other authors have pointed out that the body shape of the hybrid is more rounded than the carp and thus somewhat intermediate between the carp and goldfish. Personally, the tail looks to me like a cross between the two types of fish.
In any case, the authors provided a remarkable finding about the number of vertebrae. The goldfish had 26 vertebrae and the carp had 33-36 vertebrae. Even though the nuclear DNA came from the carp, most of the cloned hybrids had 27 or 28 vertebrae while another had 26 and another had 31. This shows that the cloned hybrid had a number of vertebrae closer to the goldfish that provided the cytoplasm than to the carp that provided the nucleus.
Moreover, all of the cloned hybrids were sterile. Sterility is common among true hybrids of different species where half of the nuclear DNA comes from one species and half from another. Here, the same results are achieved when the nuclear DNA comes from one species and the cytoplasm comes from another, suggesting a certain type of equivalence between the importance of information in nuclear DNA and information in the cytoplasm.
In general, cytoplasmic inheritance is tacitly assumed to be a function of mitochondrial DNA. This is based more on faith than on evidence, and there are a two good reasons to think it is probably not entirely true.
First, although the mitochondria contains its own DNA, most of its proteins are actually encoded by nuclear DNA, and mitochondria do not appear to export any of the proteins that they make themselves. Thus, the mitochondrial DNA might make a meaningful but perhaps relatively minor contribution to the characteristics of an organism.
Second, there is a massive amount of information contained in the cytoplasm itself that is not encoded by any genes, whether nuclear or mitochondrial, but is nevertheless inherited from generation to generation.
To understand how this is possible, let's take a look at what the inside of a cell looks like.
Image hosted by PubMed's searchable but not browsable version of the 2002 edition of Molecular Biology of the Cell
The cell is organized into a number of different compartments by a continuous system of membranes. This system of compartmentalization is absolutely essential for cellular function and for producing the biological characteristics of an organism in conjunction with the information coded in DNA.
The image is, of course, simplistic. Even within one organelle's membrane, there are further regional divisons in structure and function, and there are many fascinating localized structures that are not shown in the picture.
What is important here, though, is another point. The information needed to produce these membranous compartments is only partly coded for in DNA. Part of it resides in the membrane itself, is heritable, and is not coded for anywhere in the DNA.
Molecular Biology of the Cell, the definitive guide to mainstream molecular biology, decribes why this is so in its twelfth chapter, “Intracellular Compartments and Protein Sorting.”
Each membranous organelle is to some degree distinguished by the types of lipids it contains, but these are in turn determined by its unique set of proteins. The proteins themselves are major determinants of the oragenelle's function. These proteins have signal sequences associated with them that direct them to specific organelles.
But here's the catch! What facilitates the matching of the signal sequence to the membranous organelle for which the protein is destined? The proteins in the membrane. That's right, if there are no protein transporters in, for example, the endoplasmic reticulum, then those signal sequences that destine a protein to the endoplasmic reticulum can't be recognized by anything and have no meaning at all.
In the case of the endoplasmic reticulum, the phenomenon is even more striking, since the endoplasmic reticulum actually has to make those proteins before they can get transported anywhere anyway.
Thus, it is not just the nuclear DNA that defines the membrane, but the membrane itself. Here's the conclusion of the authors of Molecular Biology of the Cell (p. 704) in their own words:
Thus, it seems that the information required to construct an oragenlle does not reside exclusively in the DNA that specifies the organelle's proteins. Information in the form of at least one distinct protein that preexists in the organelle membrane is also required, and this information is passed from parent cell to progeny cell in the form of the organelle itself. Presumably, such information is essential for the propagation of the cell's compartmental organization, just as the information in DNA is essential for the propagation of the cell's nucleotide and amino acid sequences.
Thus, there exists absolutely critical information that is biologically heritable that is not coded for in the DNA.
To what extent do variations in this information contribute to variation between and within species?
This is a mystery that will only begin to be unraveled when more scientists drop the myopic and completely false view that organisms are largely vehicles meant as containers for self-propagating genes — a view that was always preposterous but is now more than ever completely proven false — and join the ranks of computational systems biologists who look at physiology as an integrated whole and attempt to actually ask and answer such questions.
That's not to say that most scientists actually hold such a myopic view, but scientists who study heredity essentially universally study variations in DNA sequences. The fact that we know of many examples where variations in DNA contribute to variations in heritable characteristics and do not have a similar body of knowledge about non-genetic heritable information may simply be a result of the many people asking the first type of question and almost no one asking the second type of question rather than being a result of a greater relative frequency of that type of biological inheritance.
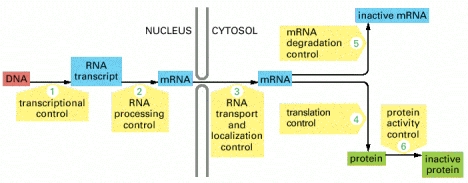
On the other hand, though we still have a great deal to learn about gene expression, we do have a very impressive body of knowledge about how cells access, utilize, control, and even in some cases restructure their genomes in response to their own needs and the needs of the organisms of which they are part. That will be the subject of the next post in this series.
Stay tuned!




thanks for sharing..
Dawkins' selfish gene is on my re-read list, its been too long since I read it and given that I know more now than I did then, I might have a different opinion now. But one way or another, I think you'll find it worth reading.
P.S. Combining your second post on De Vany's book with your 1st post, makes for an overall review that I think is mostly fair.
Dawkins' selfish gene is a must read and you are right to suppose that the title is not well chosen. He doesn't posit that the genes are selfish in themselves, what he mainly shows, is that Darwinian selection is not made on individual level (even less on species level as sometimes posited) but on another level which could be described as the gene level. The title is even worse than that because the unit he refers to is not the gene level as commonly used in biology but a "virtual gene" which can be as simple as 1 gene, but may comprise several genes.
The biggest part of his book is dedicated to explain how complex behaviours and process may spring from a group of simple units and he uses a lot of examples outside of biology to illustrate it (game theory for instance).
JA, I haven't read Pollack's book yet, but I read Ling's, which is along the same lines and I think a bit more technical. I really liked it, though I found it frustrating that he offered no explanation for what all these proteins do that he claims are unnecessary. I hope Pollack's book is better in that sense. In any case, I don't think it has much to do with what we are talking about here — whether ions require channels or pumps or not, the endoplasmic reticulum, mitochondria, golgi, and so on, exist. And their membranes contain heritable information not coded for by DNA.
Anonymous, you're welcome.
Anonymous, I made no direct reference to Dawkins because I haven't read his book yet so I don't think it's fair or wise for me to criticize him on that front. I will say, however, that if his arguments are sounder than they seem based on the title and what I have read of the basic concept elsewhere, I think he chose a rather poor metaphor to convey the concept. Genes certainly cannot be, even metaphorically, conceived of as selfish because they are themselve inert, and even if they weren't, they subjugate and in many cases sacrifice themselves to the needs of the organism. But I will put Dawkins' book on my reading list.
Thanks,
Chris
If you consider what Dawkins' classic book, The Selfish Gene, actually says, I think you'll be inclined to describe the ideas not as "preposterous", but instead as "insightful and powerful, but not the full story". The book explores non-obvious concepts that I think you'd find useful as part of your intellectual toolkit for understanding the evolved world. It's also a good read.
(And thanks for teaching me a lot, by the way!)
Thank you for you answer Chris. I may bit of an OCD type, but I'm very grateful for your answer.
Hi Chris,
Cells, Gels and the Engines of Life by Gerald Pollack, might be helpful here:
Pollack challenges the current view of the cell, the cell membrane and the role of water in the cell. Describing experiments showing that the cell can survive and function with a portion of its membrane removed, or with holes punched in the membrane, Pollack notes that the cell is really a gel, made up of cross-linked structures. And just as jello can hold almost one hundred times its weight in water—the water does not fall out of jello—so the cell holds on to its internal water by the same mechanism.
That mechanism is the structuring of water molecules along the hydrophilic surfaces of the gel matrix. Water lines up against the cell’s inner structures hydrogen-end-to-oxygen-end, not several molecules thick but dozens of molecules thick, creating a zone that excludes larger ions like sodium but not smaller ones like potassium. This selective exclusion makes the current model of complicated pumps and channels for specific compounds across the cell membrane unnecessary. The cell’s inner structure naturally excludes large ions like sodium and holds onto smaller ones like potassium.
Gels are characterized by phase transition, in which physical properties undergo abrupt transformation. Thus, with changes in pH, temperature, presence of solvents or chemical compounds, gels will suddenly expand or contract. The real mechanism is the alternating structuring and de-structuring of water inside the gels. Phase transitions of gels can explain many of the cell actions and functions such as secretion, transport, movement, muscle contraction and even cell division.
-Quoted from the review by Sally Fallon
No, …AL, I do not posit any "deus ex machina" nor any "spontaneous appearance" of organelles, and in fact I very explicitly stated that organelles cannot be synthesized de novo.
Of course it is interesting to speculate about the origin of the cell, but I was not claiming any problem for the evolvability of the modern cell. I was saying that there is heritable information contained in the organelles themselves that is not coded for in the DNA.
You seem to be trying to unravel an entirely different problem than the one I was commenting on. I do not know how the eukaryotic cell evolved — but I know something about how it currently replicates, and that replication requires pre-existing organelles in order to generate organelles in progeny cells, in the same way that the information in DNA is required for inheritance. The fact that DNA contains heritable information is, similarly, a wholly different issue from the question of how DNA came into existence.
Chris
A significant determinant of organelle membrane synthesis involves guanosine tri- & di- phosphate (GTP & GDP) energy transfering. They are mitochondrial exports functioning as messengers, so to speak.
You posit a "dues ex-machina" type of spontaneous appearance "de-novo" for organelles. I believe your mystery is resolved in light of the archaic origin of life being based on RNA. DNA based life can be considered a later refinement. And mitochondria are pressumed to be an archaic life form co-opted, or taken in, by our more complex multiple membrane enclosed life form.
Mother's line provides the mitochondrial DNA and what you say is explicitly non-DNA inherited deriviation of her cytosol. Consider that, just as "viral" RNA can "hitch" along with a membrane based life form's DNA, a cytosol RNA "hitches" along with mitochodrial DNA.
Mitochondria have sites/pores of export, you may be correct these are too small for proteins to get out. There is probably no size obstruction to the "hitching" non-mitochondrial function RNA from getting out and into the cell's cytoplasm. This may (theoreticly), in the presence of certain exported mitochondrial energy molecules, instigate the first organelle (endoplasmic reticulum?).
In other words, it's the egg that comes before the chicken…. Mitochondrial DNA has the self-assembly pattern (egg), then RNA specific to forming cytosol organelles translocates from inside the mitochondria into the cytosol (incubation), the form of the organelle as determined by RNA develops (hatching), the organelle engenders it's defining membrane and thus exists (chick), it undergoes budding and fusion processes (matures) in response to molecules/ions/energy (feed) in the cytosol to make more kinds of organelle membranes (ovulates) and perpetuate assorted organelles (lays next egg).
Response to Anonymous and …AL.
Anonymous, I think that was an overstatement. The article is five years old, and I have a lot of newer and more up to date writings on the fat-soluble vitamins. In a few months, I will comb through some of the older articles and update them where appropriate. In this particular case, however, I think the wording is a little off and failed to carry the appropriate level of uncertainty.
I would recommend my most recent article on this topic for a more up-to-date view on the science with a more appropriate language of uncertainty:
https://www.westonaprice.org/know-your-fats/2021-precious-yet-perilous.html
…AL, all proteins are encoded for by DNA. Like I said, some information needed to make the membrane is encoded by DNA, and some is not. There are no mechanisms for synthesizing membranous organelles de novo.
Chris
Organelle membrane formation involves one of the more than 60 human forms of "Rab" proteins, a type of G protein. In the cell cytosol Rab is kept linked to a guanosine di-phospate dissociation inhibitor (GDI).
Together that complex holds the program that gets Rab to the right membrane site. Once there GDI is pushed aside by a different protein, the GDI-displacement factor (GDF). Rab can get sent to the endoplasmic reticulum and other organelles.
It is my understanding that GDI, which binds to a specific Rab protein, is geneticly coded. The way that Rab complexed to GDI has it's unique determinate program set up is, I believe, still unknown.
Chris, in the article "Vitamin A On Trial: Does Vitamin A Cause Osteoporosis?" you said: "cod liver oil, in fact, because of its vitamin A content, is the only source of essential fatty acids that can lower levels of harmful, free-radical lipid peroxides, while all other sources of essential fatty acids raise lipid peroxides." Yet in the article you used as reference for this (https://www.westonaprice.org/environmental-toxins/239-dioxins-in-animal-foods-a-case-for-vegetarianism.html) you said: "Cod liver oil, on the other hand, has been shown to inhibit lipid peroxidation. One study found that cod liver oil depressed drug-induced lipid peroxidation in mice under the same conditions by which soybean oil increased lipid peroxidation.52 Another study found that feeding cod liver oil entirely abolished the increased level of lipid peroxidation found in diabetic rats.55 In both studies, the depression of lipid peroxidation was related to a sparing effect on glutathione peroxidase activity, which was also the case in rats saved from a lethal dose of dioxin by vitamin A supplementation, suggesting that the protective effect of cod liver oil is due to its high vitamin A content."
When did a "suggest" became a "sure"?